
Chromosomal Theory of Inheritance
The main role of chromosomes in heredity was first suggested in 1900 by the German geneticist Karl Correns, in one of the papers revealing the rediscovery of Mendel’s work.
Soon after, observations that similar chromosomes paired with one another throughout meiosis led directly to the chromosomal theory of inheritance, first formulated by the American Walter Sutton in 1902.
Evidence for Sutton Theory
Numerous pieces of proof supported Sutton’s theory. One was that reproduction involves the initial union of only two cells, egg and sperm. If Mendel’s model was appropriate, then these 2 gametes need to make equal hereditary contributions. Sperm, however, contain little cytoplasm, suggesting that the genetic material needs to live within the nuclei of the gametes.
Furthermore, while diploid individuals have two copies of each pair of homologous chromosomes, gametes have only one. This observation was consistent with Mendel’s design, in which diploid individuals have 2 copies of each heritable gene and gametes have one.
Lastly, chromosomes segregate during meiosis, and each set of homologue orients on the metaphase plate separate from every other set. There is however one issue with this theory.
If Mendelian characters are determined by genes located on the chromosomes, and if the independent selection of Mendelian traits reflects the independent number of chromosomes in meiosis, why does the variety of characters that type separately in a provided kind of organism often greatly exceed the variety of chromosome pairs the organism possesses? This has led many early scientists to have serious reservations about Sutton’s theory.
Thomas Hunt Morgan Experiment
In 1910 Thomas Hunt Morgan, studying the fruit fly, Drosophila melanogaster, identified a mutant male fly, one that differed noticeably from normal lies of the very same species its eyes were white instead of red.
Morgan crossed the mutant male to a normal female. All F1 progeny had red eyes. He then crossed red-eyed flies from F1 generation with each other. Of the 4252 F2 progeny Morgan took a look at, 782 (18%) had white eyes. Although the ratio of red eyes to white eyes in the F2 progeny was greater than 3:1, the results of the cross nonetheless provided clear proof that eye colour segregates.
However, there was something about the outcome that was weird and completely unpredicted by Mendel’s theory – all of the white-eyed F2 flies were male.

How could this result be explained?
Maybe it was difficult for a white-eyed female fly to exist; such individuals may not be feasible for some unidentified factor. To test this idea, Morgan test crossed the female F1 progeny with the initial white-eyed male. He got both white-eyed and red-eyed males and females in a 1:1:1:1 ratio, just as Mendelian theory anticipated. Hence a female might have white eyes. Why then existed no white-eyed females among the progeny of the original cross?
The solution to this puzzle involved sex. The gene causing the white eye trait in Drosophila resides only on the X chromosome. It is missing from the Y chromosome. A characteristic determined by a gene on the X chromosome is said to be a sex-linked trait. Knowing that the white eye trait is recessive to the red-eye trait, Morgan’s result was a natural effect of the Mendelian assortment of chromosomes.
Morgan’s experiment was one of the most essential in the history of genes because it presented the first clear evidence that the genes figuring out Mendelian characteristics do undoubtedly live on the chromosomes, as Sutton had proposed. The chromosome theory of inheritance, therefore, propounds that genes are located on chromosomes. The segregation of the white-eye trait has a one-to-one correspondence with the segregation of the X chromosome, to put it simply, Mendelian characteristics such as eye colour in Drosophila assort individually because chromosomes do.
DNA as Hereditary Material
The first proof of genetic nature of DNA was provided by a British microbiologist Frederick Griffith who made some unanticipated observations while experimenting with pathogenic bacteria. When he contaminated mice with a virulent strain of Streptococcus pneumoniae bacteria (then called Pneumococcus), the mice died of blood poisoning.
However, when he infected similar mice with a mutant strain of S. pneumoniae that did not have the virulent strain polysaccharide coat, the mice showed no ill effects. The coat was apparently necessary for virulence.
The regular pathogenic kind of this bacteria is described as the S type because it forms smooth colonies on a culture dish. The mutant types, which does not have an enzyme needed to synthesize the polysaccharide coat, is called the R kind because it forms rough colonies.

To identify whether the polysaccharide coat itself had a toxic effect. Griffith injected dead bacteria of the virulent S strain into the mice; the mice stayed completely healthy. As a control, he injected mice with a mix consisting of dead S bacteria of the virulent strain and live coatless R bacteria, each of which by itself did not harm the mice. Unexpectedly, the mice developed the illness signs and much of them passed away.
The blood of the dead mice was discovered to include high levels of live, virulent streptococcus type S bacteria, which had surface area proteins particular of the live (previously R) strains. In some way, the info specifying the polysaccharide coat had actually passed from the dead, virulent S bacteria to the live, coatless R bacteria in the mix, permanently changing the coatless R bacteria into the virulent S type.
Improvement is the transfer of genetic material from one cell to another and can modify the genetic comprise of the recipient cell.
The representative responsible for transforming Streptococcus went undiscovered until 1944. In a timeless series of experiments, Oswald Avery along with Colin Macleod andMaclyn McCarty characterized what they referred to as the “Transforming Principle”.
They first prepared a mixture of dead S Streptococcus and live R Streptococcus that Griffith had actually used. Then they removed as much of the protein as they could from their preparation, ultimately accomplishing 99.98% purity. Despite the elimination of nearly all the proteins, the transforming activity was not minimized.
Additionally, the properties of the transforming principle looked like those of DNA. The protein absorbing enzymes or RNA absorbing enzymes did not affect the transforming principles activity, however, the DNA absorbing enzyme DNase ruined all the transforming activity.
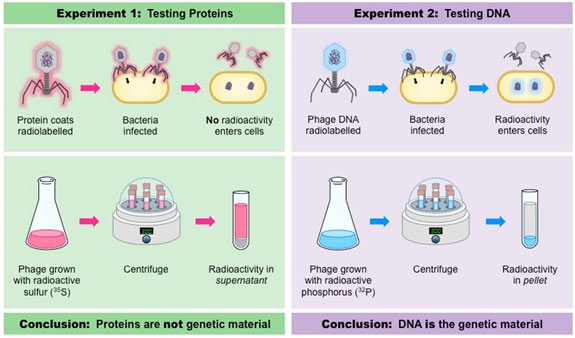
Extra evidence supporting Avery’s conclusion was provided in 1952 by Alfred Hershey and Martha Chase who explore bacteriophages T2. In some experiments, they identified viruses with radioisotope 32P, which was included in the recently synthesized DNA of grooving phage.
In other experiments, the viruses were grown on a medium including 35S, an isotope of sulphur that is incorporated into the amino acids of newly manufactured protein coats.
After the labeled viruses were allowed to contaminate bacteria, the bacterial cells were agitated strongly to remove the protein coats of the infecting viruses from the surface areas of the bacteria. This procedure eliminated nearly all of the 35S label from the bacteria. Nevertheless, the 32P label had transferred to the interior of the bacteria and was discovered in viruses consequently released from the infected bacteria. Hence, the genetic info injected into the bacteria that specified the new generation of viruses was DNA and not protein.
MCQs with Answers:
- What was suggested by Karl Correns in 1900 regarding heredity?
- a) Chromosomal Theory
- b) Mendelian Theory
- c) Genetic Variation
- d) DNA Theory
- Answer: a
- Who first formulated the Chromosomal Theory of Inheritance in 1902?
- a) Karl Correns
- b) Gregor Mendel
- c) Thomas Hunt Morgan
- d) Walter Sutton
- Answer: d
- What is one piece of evidence supporting Sutton’s theory?
- a) Diploid individuals having one copy of homologous chromosomes
- b) Diploid individuals having two copies of homologous chromosomes
- c) Diploid individuals having no chromosomes
- d) Diploid individuals having multiple copies of chromosomes
- Answer: b
- Who conducted the experiment with fruit flies and identified a sex-linked trait?
- a) Gregor Mendel
- b) Karl Correns
- c) Thomas Hunt Morgan
- d) Walter Sutton
- Answer: c
- In Morgan’s experiment, what trait did the mutant male fly exhibit?
- a) Red eyes
- b) White eyes
- c) Green eyes
- d) Blue eyes
- Answer: b
- Why were all the white-eyed F2 flies in Morgan’s experiment male?
- a) Mendelian theory was incorrect
- b) Females couldn’t have white eyes
- c) White-eyed females were not feasible
- d) Mutation occurred only in male flies
- Answer: c
- What did Morgan’s experiment provide clear evidence for?
- a) Existence of aliens
- b) Chromosomal Theory of Inheritance
- c) Mendelian assortment of traits
- d) Inheritance through environmental factors
- Answer: b
- Who provided the first evidence of the genetic nature of DNA?
- a) Oswald Avery
- b) Thomas Hunt Morgan
- c) Frederick Griffith
- d) Alfred Hershey
- Answer: c
- What bacterial strain was crucial in Frederick Griffith’s experiment?
- a) Streptococcus pneumoniae S type
- b) Streptococcus pneumoniae R type
- c) Streptococcus pneumoniae M type
- d) Streptococcus pneumoniae T type
- Answer: a
- What did Oswald Avery’s experiment identify as the “Transforming Principle”?
- a) Proteins
- b) RNA
- c) DNA
- d) Enzymes
- Answer: c
- Which isotope was used by Hershey and Chase to label the DNA in their bacteriophage experiment?
- a) 14C
- b) 32P
- c) 35S
- d) 18O
- Answer: b
- What did Hershey and Chase’s experiment confirm about the genetic material in bacteriophages?
- a) RNA is the genetic material
- b) Proteins are the genetic material
- c) DNA is the genetic material
- d) Enzymes are the genetic material
- Answer: c
- What process involves the transfer of genetic material from one cell to another?
- a) Replication
- b) Transcription
- c) Translation
- d) Transformation
- Answer: d
- What did Griffith’s experiment reveal about the transfer of genetic information between bacteria?
- a) Proteins are transferred
- b) RNA is transferred
- c) DNA is transferred
- d) Enzymes are transferred
- Answer: c
- What did the chromosome theory of inheritance propose about genes?
- a) Genes are located on chromosomes
- b) Genes are free-floating in the cell
- c) Genes are in the cytoplasm
- d) Genes are not involved in inheritance
- Answer: a
FAQs:
- What is the Chromosomal Theory of Inheritance?
- The Chromosomal Theory of Inheritance proposes that chromosomes play a central role in heredity, with genes located on chromosomes being responsible for the transmission of traits from one generation to the next.
- Who first suggested the role of chromosomes in heredity?
- The German geneticist Karl Correns first suggested the main role of chromosomes in heredity in 1900.
- When was the Chromosomal Theory of Inheritance first formulated, and by whom?
- The theory was first formulated by the American scientist Walter Sutton in 1902.
- What evidence supported Sutton’s theory?
- Evidence supporting Sutton’s theory included observations of similar chromosomes pairing during meiosis, as well as the fact that gametes have only one copy of each chromosome, which aligns with Mendel’s genetic model.
- Why did early scientists have reservations about Sutton’s theory?
- Scientists raised concerns because the number of traits appearing independently often exceeded the number of chromosome pairs in a given organism, challenging the predictions of Sutton’s theory.
- Describe Thomas Hunt Morgan’s fruit fly experiment.
- In 1910, Morgan identified a mutant male fruit fly with white eyes. Through breeding experiments, he demonstrated that the gene for eye color was located on the X chromosome, providing clear evidence for the Chromosomal Theory of Inheritance.
- Why were all the white-eyed F2 flies in Morgan’s experiment male?
- The gene for white eye color was located on the X chromosome and absent on the Y chromosome. As a result, the trait was expressed only in males.
- How did Morgan’s experiment contribute to the Chromosomal Theory?
- Morgan’s experiment provided the first clear evidence that genes determining Mendelian traits are indeed located on chromosomes, supporting Sutton’s Chromosomal Theory of Inheritance.
- What is the role of DNA as hereditary material?
- DNA serves as hereditary material by carrying genetic information. The characteristics of an organism are determined by the sequence of nucleotides in its DNA.
- What was Frederick Griffith’s contribution to understanding DNA’s hereditary nature?
- Griffith’s experiment with Streptococcus pneumoniae demonstrated the transfer of genetic material, later identified as DNA, between bacterial cells, revealing DNA’s role in heredity.
- What is the “Transforming Principle”?
- The “Transforming Principle” refers to the genetic material, identified as DNA, responsible for transforming non-virulent bacteria into a virulent form, as demonstrated in Griffith’s experiment.
- Who characterized the “Transforming Principle” and how?
- Oswald Avery, along with Colin Macleod and Maclyn McCarty, characterized the “Transforming Principle” by isolating DNA and demonstrating its ability to transform bacterial traits.
- How did Hershey and Chase provide additional evidence for DNA as genetic material?
- Hershey and Chase’s bacteriophage experiment confirmed that DNA, not protein, is the genetic material. They used radioisotopes to label DNA and protein in viruses, and only labeled DNA entered and influenced bacterial cells.
- What is the process of transformation in genetics?
- Transformation is the transfer of genetic material from one cell to another, leading to changes in the genetic makeup of the recipient cell.
- When was the Streptococcus transforming agent, later identified as DNA, discovered?
- The Streptococcus transforming agent, identified as DNA, was discovered in 1944 by Oswald Avery, Colin Macleod, and Maclyn McCarty through a series of experiments.
Wrap up
The tutorial explores the Chromosomal Theory of Inheritance, tracing its roots to Karl Correns in 1900 and the subsequent formulation by Walter Sutton in 1902. Evidence supporting Sutton’s theory includes observations on reproduction, chromosome segregation, and the pairing of homologous chromosomes during meiosis.
The tutorial delves into Thomas Hunt Morgan’s crucial fruit fly experiment in 1910, where he identified a mutant male with white eyes. The unexpected Mendelian deviations in eye color inheritance led to the discovery of sex-linked traits on the X chromosome, affirming the Chromosomal Theory of Inheritance.
Transitioning to the genetic nature of DNA, the tutorial highlights Frederick Griffith’s pioneering work with Streptococcus pneumoniae in 1928. Griffith’s experiments showcased the transformation of non-virulent bacteria into a virulent form, unraveling the role of DNA as hereditary material.
The narrative culminates with Oswald Avery, Colin Macleod, and Maclyn McCarty’s 1944 characterization of the “Transforming Principle,” identifying DNA as the genetic material responsible for transformation. Further validation came in 1952 with Hershey and Chase’s bacteriophage experiments, solidifying DNA’s role as the carrier of genetic information.
In summary, the tutorial underscores the progressive scientific journey from understanding the chromosomal basis of inheritance to recognizing DNA as the central player in heredity, marking pivotal moments in the history of genetics.